RNA-Spleißen in Menschlichen Erkrankungen
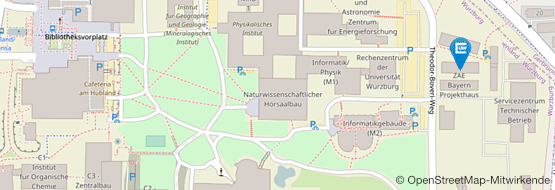
 Quantifizierung der regulierten alternativen Spleißereignisse (AS) (RNA-seq) in USP39 KO (si-USP39) HeLa-Zellen (n = 3). (B) Sashimi-Plot eines repräsentativen si-USP39-regulierten alternativen 5'-Spleißstellen-Ereignisses (n = 3). (C) Prozentsatz der annotierten 5'-Spleißstellen, die in si-USP39- und Kontroll-HeLa-Zellen verwendet werden (n = 3). (D) Spleißstellen-Stärke für kanonische (5′SS) und alternative 5'-Spleißstellen (A5′SS) von Exons mit regulären 5'-Spleißstellen (nicht reguliert) oder USP39-regulierten 5'-Spleißstellen (Wilcoxon signed-rank Test; n = 3). MaxEnt steht für die maximalen Entropie-Werte. (E) Quantifizierung der von kryptischen 5'-Spleißstellen beeinflussten RNA-Biotypen in si-USP39 HeLa-Zellen (n = 3). lncRNA, lange nicht-kodierende RNA.")
Das RNA-Spleißen ist einer der wichtigsten Regulationsmechanismen in eukaryontischen Zellen. Dereguliertes Spleißen ist die Ursache für verschiedene Pathologien, wie Retinitis pigmentosa (RP). Proben von RP-Patienten zeigen eine Verringerung der ubiquitinspezifischen Protease 39 (USP39).
In Kooperation mit Ivan Đikić an der Universität Frankfurt haben wir Zellen mit einem Knockout (KO) von USP39 analysiert, um die zugrundeliegenden Mechanismen von Spleiß-assoziierten Krankheiten zu erforschen. Diese USP39-KO-Zellen zeigen eine verstärkte Nutzung alternativer 5'-Spleißstellen, die zu einer Verlängerung oder Verkürzung des Exons vor oder hinter der kanonischen Spleißstelle führen (A,B). Die meisten der neu verwendeten Spleißstellen sind im Referenzgenom nicht annotiert (C). Dieses so genannte kryptische Spleißen wird häufig durch eine fehlerhafte Erkennung suboptimaler Spleißstellen verursacht. Tatsächlich sind die kryptischen Spleißstellen, die bei USP39 KO verwendet werden, deutlich schwächer als ihre kanonischen Gegenstücke, was auf Probleme bei der Erkennung von Spleißstellen hinweist (D). Darüber hinaus haben wir festgestellt, dass die Mehrzahl der betroffenen Gene proteinkodierend ist (E). Bei Fehlen von USP39 führt die Nutzung kryptischer Spleißstellen zur Produktion und Aggregation fehlgefalteter Proteine, was schließlich zum Zelltod führt.
Die Ergebnisse diese Projekts wurden in Pietro-Gracia et al. 2024 publiziert.
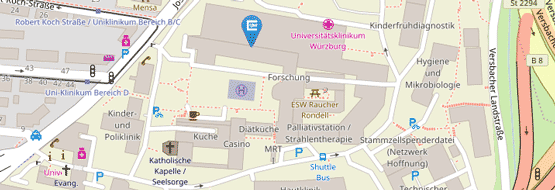
Hypoxie führt zu massiven Veränderungen im alternativen Spleißen (AS), um die Zellen an den Sauerstoffmangel anzupassen. In diesem Projekt wurde festgestellt, dass SRSF6 ein Schlüsselfaktor bei der AS-Reaktion auf Hypoxie ist. Eine starke Verringerung des SRSF6-Spiegels bei akuter Hypoxie führte zu Exon-Skipping, mit dem die Zelle das Genexpressionsprofil neu programmiert. Dieser Mechanismus ist in verschiedenen Krebsarten aktiv und erfüllt möglicherweise eine Tumorsuppressorfunktion.
In β-Zellen der Bauchspeicheldrüse fördert die Herunterregulierung von SRSF6 das Absterben von β-Zellen durch eine Dysregulation des Spleißens von zentralen Genen, welche für die Funktion und das Überleben von β-Zellen wichtig sind. In diesem Projekt haben wir die SRSF6 RNA-Bindestellen in pankreatischen β-Zellen charakterisiert und ein purinreiches SRSF6-Bindemotiv identifiziert. Wir zeigen, dass die SRSF6-Bindung an alternativen Exons das Spleißverhalten der RNAs bestimmt, da SRSF6 je nach Positionierung entweder Exon-Skipping oder Exon-Inklusion fördert. Diese Spleißveränderungen können durch den Einsatz von Antisense-Oligonukleotiden moduliert werden.
Der vollständige Artikel wurde veröffentlicht in
de Oliveira Freitas Machado et al., 2023 und Alvelos et al. 2020.
 Kreisdiagramm mit der Gesamtzahl der signifikant regulierten Lokalen Spleißvarianten (LSVs) in hypoxischen (0,2 % O2) im Vergleich zu normoxischen (21 % O2) SRSF6 überexprimierenden (OE) HeLa-Zellen, aufgeteilt in verschiedene Spleißereignisse; n = 2 Wiederholungen pro Bedingung. (B) Histogramm, das den Unterschied in der Exoninklusion (ΔPSI) in hypoxischen SRSF6 OE Zellen zeigt. Die Zahlen eingeschlossenen Exons sind oben angegeben. (C) Boxplot, der die ΔPSI-Werte von LSVs, getrennt nach mehr inkludierten, weniger inkludierten und nicht regulierten in hypoxischen WT-Zellen, in WT-Zellen nach 24 Stunden Hypoxie und die entsprechenden Werte in SRSF6 OE-Zellen zeigt. (D) Heatmap und Box-Plots zum Vergleich der LSVs in Hypoxie zwischen WT- und SRSF6 OE-Zellen. Die Cluster wurden durch k-means Clustering (k= 8) definiert. Die äußeren Violinplots und die inneren Boxplots zeigen die Verteilung der ΔPSI-Werte (Hyp-Nor) bzw. die mittleren z-Scores innerhalb jedes Clusters. (Abbildung 5 aus de Oliveira Freitas Machado et al. 2023)")